PROGRESS REPORT - YEAR 2
(September 15, 1996-September 15, 1997)
The second year we focused on processing specimens. Specimens were embedded, sectioned, and stained in order to capture images for further analysis and reconstruction. Each specimen represents an average of 160 sections. The following students were supported under this grant throughout this period of the grant: Mr. Todd Stevens (senior), Mr. Mark DeSantis (junior), Ms. Rebecca Weber (sophomore), Mr. Matthew Smatenick (junior), Mr. Peter Larson (senior), Mr. Tim Riley (junior), Ms. Robin Wilburn (junior), Ms. Mary Kate Boggiano (freshman), Mr. Charles Swart (graduate student) and Mr. Will Turner (graduate student).
Below is a report of the progress made during the second year summarized under three major sections: 1) Musculoskeletal Development, 2) Image Processing, and 3) Presentations and Publications.
Musculoskeletal Development
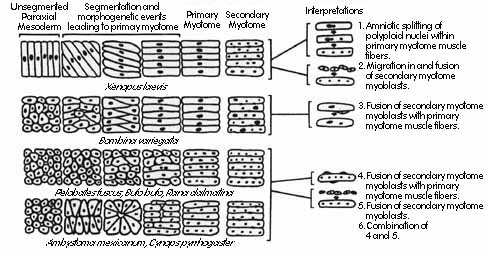
One of the goals of the project has been to search for heterochronies between different species in the patterns of myogenesis and chondrogenesis. Heterochronies may provide phylogenetic information or they may reflect historical constraints. Previous known differences in myogenesis among amphibians are summarized in Fig. 1. The few species of frogs that had been examined vary greatly in their patterns. For example, Xenopus myogenesis is precocious: myocytes express actin and myosin before myotome segmentation is complete and becomes functional before they become multinucleated (Table 1). In contrast, Bufo bufo and Rana dalmatina, primary muscle fibers become multinucleated before they become functional.
These differences may be phylogenetically useful characters. Alternatively, they may reflect historical constraints. It has been suggested, for example, that Xenopus muscle cells may become functional relatively early as part of its life history strategy of rapid development (Blackshaw and Warner, 1976; Forman and Slack, 1980). If so, then other "fast developing" anurans may share some or all the features of rapid muscle development with X. laevis, and slower developing species may have a "late" pattern of muscle development, regardless of their phylogenetic relationships.
In this context, we have compared the sequence and timing of skeletal muscle development in two species that significantly differ in developmental times. Xenopus laevis develops into free-swimming tadpoles in 3 days and metamorphoses to a juvenile frog in about 8 weeks, and Eleutherodactylus coqui a direct developer with no free-swimming tadpole that becomes a juvenile frog in about 3 weeks
Fig. 1. Patterns of amphibian myotomogenesis. Somites from left side of the animal are shown schematically as they would appear viewed from the dorsal surface, with anterior on the left. Xenopus and Bombina form functional mononucleated myocytes that become multinucleated later in development, whereas Pelobates, Bufo, and Rana become multinucleated prior to first twitch. Ambystoma and Cynops are urodele amphibians. (from (Radice et al., 1989))
Methods: To examine the sequence of events in Eleutherodactylus coqui we need to have access to recently fertilized eggs and early embryos. In mid October 1996, Gary Radice and Rafael de Sá traveled to Puerto Rico for 5 days of field work. We collected 8 adult males and 8 adult females (copies of collecting permits are attached) and transported them to our laboratory to initiate a breeding colony. A large terrarium has been constructed in de Sá's laboratory with the specific purpose of breeding Eleutherodactylus. We examined our breeding colony daily to look for eggs that had been freshly fertilized. We typically found eggs laid in clutches of approximately 15-35. The largest number of eggs in one clutch was 55. Fertility ranged from 0-100 percent, but typically was about 80%. We obtained our first clutch of eggs in December. From December to April we obtained 7 clutches of eggs, then did not find any fertile clutches during May-July, although we did find two small clutches with no fertile eggs. Because we did not obtain fertile eggs during the summer, we could not gather data on the earliest stages of myogenesis. Clutches were always laid either in the leaves of bromeliads growing in the breeding habitat, inside 2.5 cm diameter acrylic breeding tubes, or under fern leaves. When we found clutches of eggs we removed them from the breeding habitat and arranged them in a single layer on sterile filter paper inside 100 mm petri dishes. The filter paper was saturated with 10% amphibian saline to maintain a humid atmosphere, and the petri dishes were placed inside a plastic box lined with moist paper towels. The boxes were placed in the dark in an incubator maintained at 26 oC.
For standard histology, embryos were fixed in 10% neutral buffered formalin or Smith's fixative (5% w/v potassium dichromate, 4% formaldehyde, 2.5% glacial acetic acid v/v). Smith's fixative was far superior for earlier stages because it preserved the yolky tissues better. Specimens were fixed either with or without the jelly coats. Specimens were dehydrated through graded ethanols and embedded in Paraplast Plus and sectioned at 10 µm with steel knives or in glycolmethacrylate (JB4-plus) and sectioned at 0.5-1 µm with glass knives. Wax embedded tissues were stained with Milligan's Trichrome stain (potassium dichromate, acid fuchsin, phosphomolybdic acid, orange G, aniline blue, Humason, 1972). Milligan's trichrome stain yields excellent contrast between the dense orange/pink staining muscle fibers and the bluish-green collagen containing substances such as cartilage and basement membranes. Plastic embedded tissues were stained with 0.25% w/v toluidine blue in 1% w/v sodium tetraborate. Toluidine blue is metachromatic so stains nuclei blue and cytoplasm pinkish blue, making it easy to determine whether cells are multinucleate or not.
For whole mount immunocytology, we followed the protocol of (Hanken et al., 1992). Muscle cells were detected with antibody 12/101 and cartilage collagen type II was detected with antibody II-II63b (both obtained from the Developmental Studies Hybridoma Bank, University of Iowa).
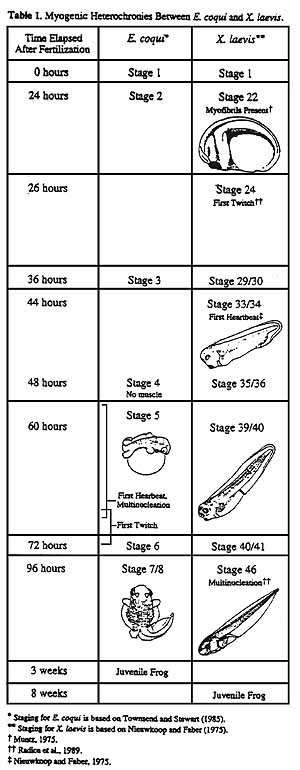
Results: Previous workers had shown that in Xenopus laevis myofibrils first appear about 15 hours post-fertilization and mononucleated muscle cells begin to twitch in axial muscles within 24 hr of fertilization, about a day before the first heartbeat.
We have found that in Eleutherodacylus coqui, myofibrils visible with light microscopy first appear during the second day of development at 26 oC. We were unable to determine how soon after myofibril formation the muscles become functional. According to Townsend and Stewart (Townsend and Stewart, 1985) first twitch occurs during day two or three, but this is only a rough estimate since the Townsend and Stewart staging series was created under field conditions and variable temperatures. Moreover, each Townsend/Stewart stage encompasses a relatively large span of development-12-24 hours-and so does not have enough resolution to distinguish priority of events with a stage. Finally, under our conditions, E. coqui seemed to develop faster than the Townsend and Stewart normal table would suggest, which has further complicated direct comparisons. Because we were unable to observe these early stages ourselves we cannot be more precise, but this should be simple matter to resolve by observing the next clutch of eggs.
In Xenopus laevis, axial muscles twitch at least a day before the heart starts beating. Our reading of Townsend and Stewart suggests that in Eleutherodactylus coqui, first heartbeat should precede skeletal muscle function, and we intend to confirm this with direct observation.
 Also, in X. laevis, myotubes become multinucleated
at about 6 days of development. In E. coqui, we have found that
myotubes become multinucleated by day two or three, probably before they
become functional, although again we as yet lack direct confirmation of
the precise time of development at which first function occurs. We have
just begun the immunocytology experiments and have not yet determined the
timing of muscle protein or cartilage collagen expression in Eleutherodactylus
coqui.. Despite these temporary uncertainties, in Table 1 we summarize
our present view of the pattern of Eleutherodactylus coqui skeletal
myogenesis compared with Xenopus laevis. We believe that E.
coqui skeletal muscle cells become functional at a later stage relative
to other developmental events than they do in X. laevis. Despite
a more rapid overall development in E. coqui to a juvenile stage,
and become multinucleated prior to function, following the Rana/Pelobates,
Bufo pattern of myogenesis rather than that of Xenopus (Figure
1).
Also, in X. laevis, myotubes become multinucleated
at about 6 days of development. In E. coqui, we have found that
myotubes become multinucleated by day two or three, probably before they
become functional, although again we as yet lack direct confirmation of
the precise time of development at which first function occurs. We have
just begun the immunocytology experiments and have not yet determined the
timing of muscle protein or cartilage collagen expression in Eleutherodactylus
coqui.. Despite these temporary uncertainties, in Table 1 we summarize
our present view of the pattern of Eleutherodactylus coqui skeletal
myogenesis compared with Xenopus laevis. We believe that E.
coqui skeletal muscle cells become functional at a later stage relative
to other developmental events than they do in X. laevis. Despite
a more rapid overall development in E. coqui to a juvenile stage,
and become multinucleated prior to function, following the Rana/Pelobates,
Bufo pattern of myogenesis rather than that of Xenopus (Figure
1).
A preliminary report of this work has been presented (see below Presentations and Publications).
The next step in our project will be to determine the timing of initial muscle myosin and cartilage collagen expression in Eleutherodactylus coqui, using whole mount immunocytology. In addition, we will be extending our analysis to embryos of a "semi-direct" developer, that is species in which the tadpoles remain within the egg for a variable period (usually about 3 weeks) before hatching. Two species common in the pet trade are possible candidates: Dendrobates auratus and Agalychnis callydrias. It may be easier to collect data on the later species because of its larger clutch size.
During the second year, we also focused on collecting whole-mounted anatomical data on the species that have been cross sectioned. This whole-mounted data strengthens the students understanding of anatomy and at the same time it helps them to understand the cross section data. The chondrocranial data collected has not been previously reported; consequently it provided an opportunity for several presentations at national meetings. In turn, this gave us the opportunity to train students in data analysis, presentation, and delivery at scientific meetings (see below Presentations and Publications).
Image Processing
From the mathematical component of the grant the worked focused on image processing. Reconstructing cartilages by means of serial sections proved to be more problematic than originally anticipated, with difficulties in registration of the sections contributing to difficulties in interpreting our 3-D images. It was decided that we would restrict attention to the muscular process; one of the largest and most visible of the chondrocranial cartilages appearing in cleared and stained whole specimens. Projections of the reconstructed muscular process onto the coordinate planes would furnish two-dimensional images for us to work with. In addition, images and illustrations of the cleared and stained whole-mounted specimens would serve to check on the effectiveness of our reconstruction protocols.
Within each coordinate plane, we wished to describe the projected image by means of its medial axis transform. Students used Mathematica to generate test images with known medial axis and known radius function. Using these images, we were able to test the skeletonization algorithms contained in several pieces of commercially available software. We were concerned both with the accuracy of the algorithm and its sensitivity to noise in the boundary of the image. As a result of these tests, we have adopted the software IPToolKit, which runs under Adobe Photoshop, for primary use in image processing and analysis.
Before an image can be skeletonized, processing is required in order to overcome boundary irregularities caused by section misalignment. Students worked with various combinations of erosions and dilations in order to smooth image boundaries and thereby reduce the complexity of the resulting skeleton. Eventually, we found a standard way to smooth the boundary using the distance map. Using statistical bootstrap methods to quantify the uncertainty we have in the image boundary remains a possible direction for our work in the coming year.
In May of 1997, with support from the NSF-CRUI grant complemented with funds from the University of Richmond's Faculty Research Committee, Dr. Kerckhove attended an Image Analysis Short Course offered by Dr. John Russ at North Carolina State University. Dr. Russ is co-creator of IPToolKit and author of The Image Processing Handbook. The short course provided a broad overview of image analysis, with time for work and discussion about our particular interests and problems. Information gained was used subsequently in working with the skeleton and distance map. We were able to write a procedure in Mathematica that takes the gray levels of pixels in the skeleton as determined by the distance map and reconstructs an image boundary based only on that data. Shape measures, which allow the original image to be described completely and for its boundary to be reconstructed uniquely, are desirable for comparative purposes. The information contained in the medial axis transform has these properties, whereas lists of shape indices do not. We have begun the task of converting shaded skeletons to numerical data.
During the summer, students worked to increase the number of images available for processing and analysis. We have produced binary stacks of various cartilages for 10 different tadpole species. Additionally, students studied stereological methods for measuring lengths and surface areas. During this study, Mark and Mary Kate became interested in the geometric problem of determining the measure of the set of hemispheres containing a given curve on the sphere. Their results, that this measure depends only on the perimeter of the convex hull of the curve, are being processed. We anticipate these result will be presented during the 97-98 year at a meeting in the form of a poster. In addition, these results will be written as a manuscript that will be submitted for publication during the fall, 1997.
In the coming year, we will proceed with the analysis of cartilage shapes. At the same time, we remain concerned about the registration problems mentioned above. At our recent Imaging Workshop, it was suggested by Dr. Stephen Pizer of UNC to employ a Bayesian approach to the medial axis transform. In this approach, sections are registered with reference to an a priori model for the cartilage shape and this may help us to overcome registration problems. This approach to processing may prove to be extremely valuable and seems well worth pursuing in conjunction with our ongoing analytical work.
Workshop
In the original project we proposed to have a workshop on digital imaging and 3D reconstruction during the first year of the grant. The delay in purchasing the computer equipment (see Annual Report Year 1) forced us to delay this workshop until the second year. The workshop took place on Saturday, September 13, 1997. All presentations will be held in Gottwald Science Center, room S-111 (deSá's lab).
Workshop Schedule
8:30 am Continental Breakfast in the Science Center E-107.
9:00-10:00 An Overview of the CRUI Project Rafael de Sá (University of Richmond) Chondrocranial Anatomy. Rafael de Sá (UR)
10:00-11:00 The chondrocranial anatomy of a primitive frog: Rhinophrynus dorsalis (Anura: Rhinophrynidae). Charles Swart (UR)
11:00-11:30 Some Technical Issues in Preparing Serial Sections Gary Radice (UR)
11:30-12:00 Heterochronies in Muscle Development. Matt Smetanick (UR)
12:00-1:00 Lunch
1:00-2:00 pm Describing the Muscular Process of the Palatoquadrate. Mike Kerckhove (UR)
2:00-3:00 Figural Representation of Shape in Image Analysis & Graphics Stephen Pizer (Keenan Professor of Computer Science, University of North Carolina, Chapel Hill)
3:00-4:00 Imaging Structures in the Brain Princy Susan Quadros (UR)
4:00-5:00 Seven Years of Teaching with Digital Microscopy Robert Blystone (Professor of Biology, Trinity University, San Antonio TX)
5:00-8:00 Reception at the Deanery.
Presentations and Publications
During the second year we presented several papers and submitted papers for publication.
Presentations:
· R. O. de Sá. 1997. Anatomical characteristics of Phyllomedusinae tadpoles. Third World Congress of Herpetology. Abstract and Oral Presentation. August, 1997.
· R. O. de Sá and E. O. Lavilla. 1996. Descripción de la larva de Pseudis minuta (Anura: Pseudidae) y de su morfología oral interna. IV Latinamerican Congress of Herpetology, Santiago de Chile, Chile, October 1996.
The following student presentations were at the Combined meetings American Society Ichthyologists and Herpetologist, Herpetological League, and Society for the Study of Amphibians and Reptiles. Seattle Washington, June 1997:
· P. Larson and R. O. de Sá. Chondrocranial morphology and skeletogenesis in the genus Leptodactylus (Amphibia: Anura: Hylidae). Oral Presentation.
· C. Swart and R. O. de Sá. The chondrocranium of Rhinophrynus dorsalis. Poster Presentation
· G. Schwartz, T. Stevens, R. O. de Sá. Chondrocranial anatomy, skeletogenesis, and internal oral morphology of three species of Ptychohyla (Amphibia: Anura: Hylidae). Poster Presentation.
· R. Weber and R. O. de Sá. Diagnostic characteristics of Hypopachus tadpoles (Anura: Hylidae). Poster Presentation.
· T. D. Riley and R. O. de Sá. Chondrocranial anatomy and skeletogenesis in two species of neotropical Hyla. Poster Presentation.
· M. Smetanick, G. P. Radice, and R. O. de Sá. Patterns of musculoskeletal development in the direct and indirect developing anurans. Poster Presentation.
The following students presented their research at the Undergraduate Research Symposium, University of Richmond, April 1997.
· R. Wilburn, M. DeSantis, and M. Kerckhove. Biological shape: Math and the tadpole chondrocranium. Abstract and Poster Presentation
· G. Schwartz, T. Stevens, R. O. de Sá. Chondrocranial anatomy, skeletogenesis, and internal oral morphology of three species of Ptychohyla (Amphibia: Anura: Hylidae). Abstract and Poster Presentation
· R. Weber and R. O. de Sá. Diagnostic characteristics of Hypopachus tadpoles (Anura: Hylidae). Abstract and Poster Presentation
· P. Larson and R. O. de Sá. Chondrocranial morphology and skeletogenesis in the genus Leptodactylus (Amphibia: Anura: Hylidae). Abstract and Oral Presentation.
· M. Smetanick and G. Radice. Patterns of Musculoskeletal Development in Direct and Indirect Developing Anurans. Abstract and Oral Presentation.
Publications:
· R. O. de Sá and E. O. Lavilla. 1997. An apparent case of heterochrony, the tadpole of Pseudis minuta (Anura: Pseudidae). Amphibia-Reptilia 18:229-240
· R. O. de Sá and Shannon Hill. 1997. Chondrocranial anatomy and skeletogenesis in Dendrobates auratus (Anura: Dendrobatidae). Accepted for publication in: Journal of Herpetology.
· E.O. Lavilla and R. O. de Sá. 1997. Estructura del condrocraneo y esqueleto visceral de larvas de Pseudis minuta (Anura: Pseudidae). Submitted to: Alytes.
· C. Swart and R. O. de Sá. 1997. The chondrocranium of the morphology Mexican Burrowing Toad Rhinophrynus dorsalis (Anura: Rhinophrynidae). Submitted to: Journal of Herpetology.
· A. T. d'Heursel and R. O. de Sá. 1997. Comparing the tadpoles of Hyla geographica and Hyla semilineata (Anura: Hylidae). Submitted to: Journal of Herpetology.
Current and Pending Support of Senior Personnel
During the 1996-1997 academic year, Dr. Gary Radice submitted a grant proposal entitled "Origin and Identity of Amphibian Lymph Heart Muscle." This proposal was not funded and has not been resubmitted.
Dr. Rafael de Sá has submitted a proposal to the NSF-RUI program entitled "Evolutionary relationships within the Neotropical frog genus Leptodactylus." Funding of this proposal is pending. If funded, starting date for this projects has been proposed for May 1, 1998. The pending proposal does not include salary during the summer of 1998 and includes a commitment for Dr. de Sá of 2.5 months during the academic year. Dr. de Sá's commitment under the C-RUI is equivalent to 3.35 months of the calendar year. Furthermore, the C-RUI will end in September 1998, overlapping with the pending proposal only for only 4 months.
Literature Cited
Last modified January 9, 1998.
Return Home